
Testo del 24.04.2014
Vorrei tentare di riunire un piccolo gruppo di appassionati per sviluppare una ricerca sui sistemi di comunicazione nell’arnia per comprendere più a fondo le reti neurali che uniscono i singoli soggetti dell’alveare concretando un pensiero comune e condiviso. Lo studio potrebbe poi svilupparsi in due direzioni differenti dagli interessanti risvolti pratici: da una parte le reti neurali biologiche che possono essere oggetto di approfondimento per lo studio della “telepatia umana” e del sistema di comunicazione tra cellule e dall’altra le reti neurali artificiali che possono essere oggetto di approfondimento per i sistemi informatici e di telecomunicazione futuri (vedi computer biologici). Se siete interessati potete farvi un’idea di massima leggendo questa voce di Wikipedia:
Rete neurale
INSETTI SOCIALI (nella Treccani)
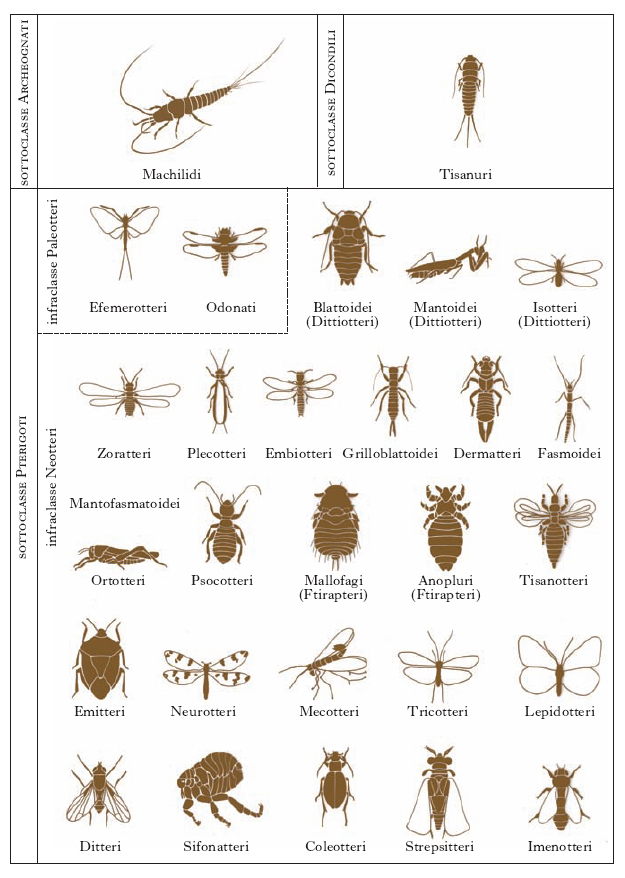
insètti sociali Specie di insetti (detti anche eusociali) che vivono in società organizzate, suddivise in caste, nelle quali i singoli individui agiscono in funzione delle necessità collettive. Presentano tre caratteristiche: la prole originata dagli individui riproduttori viene allevata da più individui in modo cooperativo; si registra la presenza di una casta sterile all’interno della specie; nel medesimo periodo coesistono all’interno della colonia gli individui riproduttori, la prole adulta e la prole immatura. Questo tipo di società si riscontra negli Imenotteri (formiche, api, vespe), negli Isotteri (termiti) e negli Omotteri (afidi).
INSETTI SOCIALI – APPROFONDIMENTO
di Stefano Turillazzi e Mary Jane West-Eberhard (Enciclopedia della Scienza e della Tecnica)
EVOLUZIONE DELLA SOCIALITÀ – GLI INSETTI E IL COMPORTAMENTO SOCIALE
Un gruppo sociale non è semplicemente un insieme di individui solitari che si ritrovano insieme in particolari circostanze. Il comportamento sociale propriamente detto è tipico di quegli organismi che vivono in gruppi all’interno dei quali le relazioni tra i singoli individui sono caratterizzate da schemi ben precisi e talvolta di elevata complessità e organizzazione. In questi gruppi si distinguono ruoli e compiti che mutano profondamente il comportamento dei diversi individui rispetto a quello di specie simili ma solitarie. Da queste premesse si rileva come la vita sociale sia caratteristica di un numero limitato di specie tra cui, ovviamente, la nostra. A partire dagli anni Settanta del Novecento lo studio di questo particolare fenomeno ha fatto sì che l’elenco delle specie sociali si allungasse in modo considerevole. In gran parte ciò si deve alla grande quantità di ricerche scaturite dalle nuove idee evolutive, portate avanti da numerosi ricercatori, e in particolare ai fondamenti della teoria nota come ‘selezione di parentela’ (kin selection). Il libro Sociobiology: the new synthesis di Edward O. Wilson (1975) ha sancito la nascita di questa nuova scienza che studia l’evoluzione del comportamento sociale, dando il via a una serie di studi approfonditi sulla storia naturale, sull’eco-etologia e sull’evoluzione delle specie sociali. In questa storia gli Insetti sociali hanno un posto importante. È indubbio, innanzi tutto, che questo fervore di studi e ricerche è iniziato per rispondere a un preciso interrogativo, che Darwin stesso si era posto in On the origin of species, sull’origine e l’evoluzione delle caste sterili negli Insetti sociali.L’eusocialità e gli ordini socialiI criteri utilizzati dagli scienziati per definire il comportamento sociale, propriamente detto ‘eusocialità’, sono essenzialmente tre: la presenza nel gruppo di un allevamento in comune della prole; l’esistenza di una sovrapposizione di generazioni di individui abili al lavoro; infine, cosa più importante, la presenza di una differenziazione dei ruoli all’interno del gruppo, con individui che si riproducono e individui che non lo fanno e che si specializzano in compiti legati all’allevamento della prole e al mantenimento o alla difesa della colonia e del luogo dove questa si è instaurata. Una simile organizzazione di gruppo può essere trovata, temporaneamente, anche in molti Vertebrati, ma esempi in cui la sterilità di una parte dei componenti del gruppo duri per tutta la vita e si trovi unita a una diversa morfologia delle due classi di individui sono stati finora descritti solo in alcuni invertebrati e in due specie di Vertebrati Roditori. Uno di questi è Heterocephalus glaber, roditore dell’Africa orientale che costruisce grandi nidi sotterranei e che presenta colonie molto simili a quelle delle termiti. Gli Insetti presentano il numero più consistente di specie in cui è stata accertata l’eusocialità. Gli ordini in cui sono state ritrovate specie eusociali sono cinque: gli Isotteri (termiti), gli Emitteri Omotteri (afidi), i Tisanotteri, i Coleotteri e gli Imenotteri (api, vespe e formiche); quest’ultimo è l’ordine in cui la socialità, nel corso dell’evoluzione, si è sviluppata più volte e indipendentemente. Tra gli Imenotteri, alcune famiglie, come quella dei Formicidi, le formiche, presentano specie esclusivamente sociali. Mentre tutti gli Isotteri, le termiti, sono eusociali con vari gradi di eusocialità, ben più rari sono gli esempi di eusocialità negli altri tre ordini. L’evoluzione dell’eusocialitàCome può spiegarsi un comportamento altruistico, cioè un comportamento che, abbassando la probabilità di sopravvivenza e di riproduzione dell’operaia (l’individuo che compie l’atto), aumenti contemporaneamente le probabilità di sopravvivenza e di riproduzione della regina (l’individuo verso cui l’atto è rivolto)? William D. Hamilton negli anni Sessanta del Novecento esaminò il fenomeno delle caste sterili degli Insetti sociali e in particolare degli Imenotteri, mettendo in relazione l’eusocialità con il meccanismo di riproduzione e di determinazione del sesso proprio di questo ordine di Insetti: quello dell’aplodiploidia. Negli Imenotteri le femmine si sviluppano, infatti, da uova fecondate, diploidi, mentre tutti i maschi prendono origine da uova partenogenetiche, aploidi, e producono spermatozoi uguali dal punto di vista genetico. Questo causa asimmetrie nei coefficienti di parentela: due sorelle hanno infatti in comune, in media, 3/4 dei loro geni, mentre la parentela tra madre e figlie è solo di 1/2. Hamilton suggerì che il comportamento delle operaie volto a favorire la produzione di sorelle, attraverso la riproduzione della regina madre, fosse selezionato positivamente perché, in questo modo, la percentuale dei loro geni trasmessi ad altri individui sarebbe stata più alta di quella che esse avrebbero potuto raggiungere riproducendosi direttamente. Sebbene la selezione di parentela abbia avuto sicuramente importanza nell’origine e nell’evoluzione degli Insetti sociali in generale, non sembra che essa sia stata l’unica forza selettiva in gioco. Infatti la condizione di parentela di 3/4 può essere ottenuta raramente, specialmente nelle specie socialmente primitive, a causa della frequenza di accoppiamenti multipli e della presenza di più regine nelle colonie. Inoltre alcuni Insetti sociali (per es., le termiti) non sono aplodiploidi. L’eusocialità si è quindi probabilmente originata ed evoluta tanto attraverso la selezione individuale che attraverso la selezione di parentela; si pensa che quest’ultima abbia agito in particolare su gruppi familiari, secondo la teoria della famiglia poliginica. Le società delle vespe: dal semplice al complessoLe colonie di vespe e formiche presentano organizzazioni sociali assai complesse, con sistemi di comunicazione basati su feromoni e con una casta operaia specializzata anche per lavori di vario tipo: difesa, raccolta del nutrimento, cura delle larve, ecc. Invece tra le vespe e le api troviamo, accanto a specie estremamente evolute dal punto di vista sociale, altre che soddisfano i tre criteri di eusocialità solo in alcune fasi del loro ciclo coloniale e in cui le femmine mostrano una flessibilità, estesa alla vita adulta, nella possibilità di appartenere a una data casta. Api e vespe rappresentano quindi un oggetto di studio molto importante, perché il confronto permette di ricostruire la probabile storia evolutiva del comportamento sociale. La famiglia dei Vespidi è composta da sei sottofamiglie: Euparagine, Masarine, Eumenine, Stenogastrine, Polistine, Vespine; le ultime tre presentano fenomeni di socialità a vario livello.
LA COSTRUZIONE DEL NIDO
Il nido rappresenta un importante fattore per l’origine e l’evoluzione della vita di gruppo negli Insetti. Esso conferisce al suo costruttore una sorta di controllo sull’ambiente; con la creazione di un ambiente più stabile, rende la fondazione di nuove colonie più difficile e limita l’abbandono della colonia da parte dei giovani, con il risultato di far aumentare le dimensioni della stessa e di rendere più complesse le interazioni tra gli individui. Al tempo stesso, il nido agisce in molti casi come protezione nei confronti di predatori e parassiti, rendendo possibili alcune particolari forme di comunicazione tra gli individui che compongono la colonia e migliorando la capacità di allevamento della prole grazie alla possibilità di immagazzinare cibo; esso pone però anche alcune limitazioni al suo costruttore in ragione del vincolo di dipendenza che instaura.L’architettura del nido nelle vespe sociali rappresenta un campo di studio di enorme interesse, perché si può senz’altro affermare che proprio in questi Insetti sociali essa raggiunge la sua estrema variabilità e complessità. L’architettura del nido è il risultato di varie pressioni selettive: la prole delle vespe sociali è estremamente vulnerabile a causa della limitazione nei movimenti – larve apode e costrette in celle a misura – e per il grande numero di larve stivate assieme, che rende il nido una fonte di cibo molto ambita per predatori Vertebrati e invertebrati. Queste caratteristiche sono probabilmente il risultato secondario della vita in un nido, in quanto un nido costruito per proteggere la prole rende la prole stessa più vulnerabile e ciò a sua volta aumenta l’importanza del nido, in una sorta di processo autorinforzante.La prole immatura deve necessariamente contare sulla difesa degli adulti per la sua sopravvivenza. Questa difesa può essere ‘diretta’, e questo spiega la dotazione negli adulti di armi temibili, come pungiglioni, secrezioni velenose e urticanti, o ‘indiretta’, mediante strutture protettive fornite dal nido stesso, quali il peduncolo e l’involucro. Il peduncolo è una struttura che riduce l’accesso ai predatori terrestri, quali le formiche, alle celle in cui sono allevate le larve e custodite le riserve di cibo. L’involucro ha la stessa funzione difensiva, ma è efficace anche nei confronti di predatori non terrestri e di parassiti. Tutte e due le strutture comportano però una quantità di materiale per la costruzione del nido molto maggiore rispetto a quella necessaria per la realizzazione di semplici cellette di covata. La quantità di energia spesa per la costruzione di un nido complesso va quindi messa nel conto delle forze selettive che hanno modellato l’architettura del nido stesso. Talvolta la costruzione di un nido con un’architettura particolarmente adattata a camuffarsi con l’ambiente, e quindi a conferire una difesa maggiore alle larve, comporta la riduzione delle dimensioni del nido stesso e, di conseguenza, una sorta di involuzione del livello di socialità di una particolare specie. Anche la scelta e la costituzione del materiale usato per la costruzione del nido influenzano l’architettura e, indirettamente, la socialità dei costruttori: per esempio, le Stenogastrine fanno uso di fibre legnose molto corte, impastate con saliva priva però di adeguate sostanze in grado di tenere insieme le fibre stesse in modo ottimale. Le Polistine sono invece capaci di utilizzare fibre vegetali più lunghe e di tenerle assieme con sostanze assai più efficaci; ciò permetterebbe loro di costruire nidi molto più grandi e, di conseguenza, di dar vita a colonie più numerose. Il nido può essere inoltre estremamente importante per fornire indicazioni di carattere filogenetico e, al limite, per distinguere specie diverse ma morfologicamente simili. Le vespe si dimostrano quindi un eccellente oggetto di studio per l’evoluzione del comportamento di costruzione.
L’ORIGINE DEL DUALISMO REGINA-OPERAIA
La divisione in caste è direttamente connessa al problema dell’altruismo. L’evoluzione dell’eusocialità ha due fasi principali: l’origine dell’altruismo e l’origine dei tratti che determinano il completo e irreversibile differenziamento riproduttivo degli individui. Anche in questo caso le vespe si dimostrano buoni modelli di studio, perché esiste una grande varietà di modalità con cui il differenziamento riproduttivo avviene, a livello comportamentale, anatomo-fisiologico o morfologico. È il genere Polistes il miglior soggetto per studiare le caratteristiche degli individui sterili e valutarne i costi e i benefici del comportamento. In effetti l’origine e l’evoluzione di questi tratti rappresentano la principale problematica sulla quale si incentrano gli studi sociobiologici. Come si è originato il dualismo regina-operaia? Il fenotipo operaio si è probabilmente originato dalla condizione ovarica delle vespe solitarie, dove con ‘condizione ovarica’ si intende tutto quel particolare sistema in cui le dimensioni degli ovari sono legate all’attività ormonale (principalmente quella dell’ormone giovanile, che ha notoriamente effetto gonadotropo negli Insetti adulti) e il comportamento viene influenzato da esse tramite coordinazioni neurormonali. Nelle vespe solitarie esiste un ciclo ovarico ben preciso, che corrisponde a un ciclo comportamentale in cui si succedono ripetitivamente la costruzione di una cella, quando l’uovo si avvicina a essere deposto e, nella fase che segue la deposizione e che coincide con un ovario privo di uova sviluppate, comportamenti di guardia e di approvvigionamento della larva. Durante la fase di ovario sviluppato, inoltre, le femmine di molte specie solitarie mostrano una marcata aggressività verso conspecifici in vicinanza del loro nido. Si può quindi concludere che anche la dominazione aggressiva che si riscontra nelle specie sociali come Polistes, uno dei meccanismi fondamentali della loro organizzazione sociale, rappresenta probabilmente un tratto ancestrale ed era già presente nei progenitori solitari delle vespe sociali. Si è anche osservato che, se una femmina non è in grado di deporre un uovo sviluppato per la mancanza di un luogo adatto o per carenza di nutrimento, o ancora perché impedita da un conspecifico, l’uovo può essere riassorbito lasciando una traccia nell’ovario. La divisione riproduttiva del lavoro nelle vespe sociali sembrerebbe dunque correlata all’aggressività legata alle condizioni ovariche, causando lo sdoppiarsi del ciclo ovarico proprio delle vespe solitarie in una fase regale, caratterizzata da uova sviluppate, e in una fase operaia, caratterizzata da uova non sviluppate, con la concomitante divisione dei ruoli comportamentali.Tra le Stenogastrine, in varie specie delle quali è stata descritta la presenza di gerarchie di dominazione tra le femmine che formano le colonie, le femmine mostrano una plasticità comportamentale molto ampia, con varie strategie alternative che vanno dalla fondazione e riproduzione solitarie all’aggregazione o all’usurpazione di colonie di parenti, inclusa quella materna, e di non parenti. Da questo punto di vista è chiaro che il grado di socialità nelle popolazioni di una specie e il ruolo di un individuo in una colonia sono influenzati da contingenze determinate dall’ambiente sociale in cui esse si vengono a trovare. Lo stesso è vero per tutte quelle vespe primitivamente eusociali, nelle quali il ruolo di una femmina all’interno di una colonia è determinato dal ciclo della colonia stessa e dalla presenza di regine attive, ma dove sono possibili inversioni dei ruoli sociali. Un chiaro esempio di questo è rappresentato dalle operaie o femmine subordinate che acquistano la capacità di riprodursi se la regina viene a mancare. Anche nelle vespe che presentano una differenziazione in caste più netta, la condizione ovarica regola nel tempo la varietà del comportamento degli individui, contribuendo in modo determinante alle complesse organizzazioni sociali delle colonie di questi Insetti. Tutto ciò mostra come le tre fasi principali del ciclo ovarico delle vespe solitarie possano essere servite come base per l’evoluzione della divisione del lavoro e delle diverse modalità che lo caratterizzano nella socialità delle vespe.
IL CONTROLLO DELLA COLONIA
Il complesso dei comportamenti ritualizzati, che si sostituiscono alle lotte e si identificano nell’imposizione dell’individuo dominante e nell’acinesi di quello subordinato, ha l’effetto non solo di determinare il ruolo di riproduttrice ma anche quello di organizzare la divisione del lavoro, con le femmine subordinate che si assumono il ruolo di operaie. Il meccanismo di controllo della colonia sarebbe quindi, nel periodo della fondazione, principalmente comportamentale e dovuto all’aggressività della femmina dominante nei confronti delle sue subordinate. Il riconoscimento dei ruoli tra gli individui che compongono la colonia è alla base della ritualizzazione dei moduli comportamentali che si sostituiscono alle lotte, impedendone un’intensificazione ed evitando disastrosi confronti una volta che la gerarchia sia stata stabilita. Il rango è direttamente correlato alle dimensioni ovariche dei vari individui, nonché all’attività di ghiandole endocrine, quali i corpora allata (responsabili della produzione dell’ormone giovanile). Il comportamento di dominazione è inoltre influenzato da parecchi fattori endocrini e la deficienza di un sistema può essere compensata dalla maggiore attività di un altro. Questi fattori endocrini sono dati dall’attività non necessariamente sinergica dei corpora allata e degli ovari; in alcune specie potrebbero essere coinvolte anche prostaglandine liberate in seguito all’accoppiamento. Le differenze fisiologiche possono essere sia geneticamente controllate sia indotte dal nutrimento larvale, o anche essere influenzate dalla data di sfarfallamento o da altri eventi avvenuti prima dell’ibernazione. All’interno dei gruppi di fondazione, altri attributi, quali le dimensioni e l’ordine di arrivo al nido, possono comunque influenzare l’organizzazione della gerarchia.Riguardo ai meccanismi che causano la progressiva perdita di fertilità delle subordinate, essi sono legati alla presenza sul nido delle femmine dominanti e al loro comportamento; quest’ultimo, tuttavia, da solo è insufficiente a inibire completamente la capacità di deposizione delle femmine ausiliarie. Diversi fattori possono quindi agire contemporaneamente per sterilizzare le subordinate e tra questi è importante il fatto che la femmina dominante, detta ‘alfa’, sia perfettamente ovificante. Fenomeni di dominazione sono alla base dell’organizzazione sociale delle colonie anche dopo la nascita delle operaie. Queste attraversano un ciclo di sviluppo ovarico prestabilito, che ha il suo massimo verso il decimo giorno dallo sfarfallamento. In generale questo fenomeno determina l’insorgenza di individui che possono, in determinate condizioni coloniali, come la decadenza della femmina dominante, iniziare a deporre. In Polistes, a seconda della specie, le interazioni tra le femmine sono caratterizzate da fortissima aggressività e dispotismo, fino ad arrivare a un’assenza pressoché totale di ostilità a cui corrisponde una divisione del lavoro appena accennata. Gerarchie di dominazione sono state individuate e descritte anche in altre vespe. Nelle piccole società delle Stenogastrine, per esempio, queste sono presenti in diverse specie. Gerarchie di dominazione sono ben evidenti, anche se con varie sfumature del comportamento aggressivo, nelle Polistine caratterizzate da colonie simili a quelle di Polistes, come Belonogaster, Ropalidia, Mischocyttarus, Parapolybia.Vi è inoltre il problema del riconoscimento individuale e quindi dei meccanismi di controllo usati dalla regina all’interno di colonie molto numerose. Il controllo della colonia è stabilito e mantenuto con contatti fisici diretti e aggressivi e con l’oofagia e, in un momento successivo, la superiorità aggressiva si connette a segni di riconoscimento individuali prontamente identificabili dalle compagne di nido. Questo permetterebbe agli individui subordinati di evitare i costi di ulteriori confronti. In ultima istanza l’interazione diretta diverrebbe sempre più rara e sempre più sostituita da un segnale della presenza della regina. Questi segnali, evidentemente di origine chimica, portano alla formazione di corti regali, dove la regina è costantemente circondata da un gran numero di operaie che la seguono, la nutrono e la accudiscono. Con l’avvento del segnale di riconoscimento chimico emesso dalle regine, che agisce come una dichiarazione di rango e di stato, il passaggio da sostanze con un effetto di riconoscimento (releaser) a sostanze con un effetto di inibizione (primer) ha rappresentato un passo molto breve nell’evoluzione. Questo non vuole, tuttavia, significare che i fenomeni di competizione siano scomparsi completamente e che le colonie siano utopistiche, pacifiche società. Il concetto di ‘dominazione’, limitato al concetto ristretto dell’aggressione e dell’attacco fisico, può essere allargato fino a comprendere anche le più sottili e non violente interazioni tra le operaie delle vespe sociali più evolute. Si troverebbe così che questo sistema di dominazione coinvolge virtualmente tutti gli individui di una colonia. In realtà si osserva che operaie di tutte le specie di vespe sociali possono riprodursi e possono sviluppare e deporre uova almeno in un certo momento della loro vita riproduttiva. Inoltre, la presenza di una regina attiva è necessaria per inibire lo sviluppo ovarico e un comportamento di tipo regale nelle operaie. In altre parole, la colonia si trova sempre sotto una costante tensione competitiva, mantenuta dal vertice della gerarchia riproduttiva.
PARASSITISMO E RICONOSCIMENTO
Un fenomeno che potrebbe aiutarci a chiarire l’effettiva natura dei meccanismi del controllo coloniale in Polistes è quello dei parassiti sociali. Un parassita sociale è una specie che ha perduto la casta operaia e le cui femmine, per riprodursi, devono usurpare il nido di un’altra specie e utilizzarne le operaie per l’allevamento della propria prole. Conosciuti in tutti gli Imenotteri sociali, questi particolari parassiti si ritrovano anche nelle Vespine. L’effetto che i parassiti hanno sulla fitness degli ospiti è, in taluni casi, catastrofico e tutti gli individui riproduttori che la colonia produce appartengono alla specie usurpatrice. In molte colonie, tuttavia, il parassita non raggiunge il controllo della colonia, con conseguente sviluppo degli ovari e deposizione di uova da parte delle operaie del nido occupato e, in certi casi, con la fondazione di altri nidi da parte di individui che abbandonano il nido materno. L’invasione del nido ospite da parte delle femmine parassite avviene sempre in un particolare momento del ciclo coloniale, quando ancora le operaie non sono sfarfallate o sono presenti solo in piccolo numero. Studi sulle strategie di invasione del nido indicano che Polistes parassiti usano due principali tattiche per prendere possesso del nido ospite: una aggressiva e l’altra non aggressiva. Nella prima, il parassita combatte accanitamente contro le fondatrici ospiti e scaccia dal nido le femmine dominanti dopo averle spesso mutilate delle zampe, mentre mantiene le subordinate nel nido sottoponendole a un intenso trattamento che ricorda da vicino quello di dominazione. Un’altra specie parassita raggiunge invece il suo scopo in un tempo più lungo, sopportando senza reagire gli attacchi sempre meno convinti della fondatrice ospite, fino a imporsi gradualmente come dominante. Se la fase di conquista del nido è molto rischiosa, il mantenimento del controllo della colonia rappresenta per la regina della specie parassita un problema molto più difficile da risolvere: essa deve indurre individui completamente estranei, le operaie, a lavorare per lei, rinunciando a riprodursi e perdendo anche qualsiasi possibilità di allevare prole imparentata. Questa considerazione porta a pensare che le regine delle specie parassite impieghino mezzi di controllo della colonia molto più potenti di quelli usati dalle regine di Polistes e che non si limitano alla sola dominazione comportamentale. Si possono avanzare due ipotesi: la prima contempla la possibilità che i parassiti impieghino feromoni inibitori dell’aggressività, trasferiti agli ospiti durante le accurate operazioni di leccatura effettuate sul loro corpo nel corso delle dominazioni. Nei confronti della prole immatura i parassiti potrebbero essere inoltre capaci di manipolare o altrimenti di camuffarsi con l’odore della colonia ospite e di ingannare le future operaie sulla loro effettiva parentela con le usurpatrici. Con ‘odore di colonia’ si indica quel particolare segnale che permette a ciascun membro di discriminare tra compagni di nido e individui estranei. In vari Insetti sociali, tra cui Polistes, questo segnale è di natura chimica ed è costituito da una miscela di idrocarburi a elevata massa molecolare che ricopre la cuticola dei vari individui. Questa impronta chimica è molto simile per i membri di una stessa colonia e, nel caso di Polistes, è presente anche sul materiale del nido; essa viene appresa dagli adulti durante un breve periodo sensibile subito dopo lo sfarfallamento.Una volta preso possesso della colonia, le femmine parassite compiono un’accurata strusciatura del nido, che può durare anche varie ore, con gli sterniti addominali. Un simile comportamento, per la verità molto evidente, è riportato anche per quei parassiti sociali che attaccano le colonie di alcune specie di Vespini e sembra legato all’applicazione sulla superficie del nido di secrezioni prodotte da ghiandole addominali o a un’attiva modifica dell’impronta idrocarburica da parte della regina parassita, capace di assumere l’odore della colonia ospite. L’osservazione accurata di alcuni comportamenti e la riconsiderazione di un fenomeno ampiamente diffuso in alcune specie di Polistes, quello dell’usurpazione intraspecifica del nido, hanno messo tuttavia in evidenza che questi comportamenti, così evidenti nei parassiti sociali, sono presenti, seppur molto più sfumati, anche nel repertorio degli ospiti. I parassiti sociali, pertanto, si mostrano soggetti di studio molto interessanti, perché possono essere considerati amplificatori dei sistemi di controllo coloniale presenti nelle vespe ospiti.
FONTE
—
Insetti sociali
Gli Insetti sociali appartengono a due ordini; il più antico è quello degli Isotteri (Térmiti) interamente sociale, il secondo quello degli Imenotteri, che comprende Vespe, Api e Formiche che presentano anche forme sociali.
Vi è una differenza importante tra le generiche aggregazioni di insetti e le società. Le aggregazioni sono dovute alla convergenza verso un comune bisogno. Nelle aggregazioni talvolta gli individui cominciano a svelare una certa interattrazione, e pertanto si parla di gregarietà, che può essere semplice oppure di tipo coordinato. Nel primo caso, gli individui, pur mantenendosi in contatto per effetto di interattrazione, manifestano indipendenza di movimenti; nel secondo caso, gli individui compiono azioni coordinate, come accade, ad esempio, nello spostamento in massa di un’orda di cavallette.
Nell’ape, la regina è l’unica femmina fertile dell’alveare. Le altre femmine, dette operaie, eseguono tutti i lavori necessari per la sopravvivenza della comunità: cura delle larve e della regina, costruzione e riparazione dell’alveare, raccolta di nettare e polline (api bottinatrici), produzione di miele, cera e pappa reale, difesa. La regina, più grossa, ha anche una vita più lunga di quella delle operaie (alcuni anni anziché pochi mesi). La sua unica occupazione è la deposizione delle uova. Essa produce un feromone che inibisce lo sviluppo degli organi riproduttori delle operaie. In caso di morte della regina, e se non esistono uova con età inferiore a tre giorni, una delle operaie sviluppa gli organi riproduttori e tenta di sostituire la regina anche se da lei nasceranno solo fuchi perché non è stata fecondata. Per quasi tutto l’anno, la regina depone uova da cui si ottengono operaie. Nella stagione adatta, alcune uova di sesso femminile sono poste in celle più grandi, le celle reali, e le larve che ne nascono sono nutrite con pappa reale. Nel frattempo vengono deposte uova che produrranno i maschi, le cui larve si sviluppano in celle apposite. La regina che sfarfalla per prima uccide le larve “concorrenti” e prende il posto della vecchia regina. Quest’ultima, con parte delle operaie, sciama per costruire un nuovo alveare. La nuova regina esegue il volo nuziale, con i maschi con cui si accoppia. Essa accumula gli spermatozoi in un organo apposito, che li mantiene vitali e le permette di fecondare le uova che man mano produce nel corso della sua vita. I maschi vengono abbandonati.
Le api sono insetti evoluti, olometaboli e con appendici specializzate, come il pungiglione addominale. Hanno un comportamento complesso, in cui spicca la danza con la quale un’ape bottinatrice comunica alle compagne direzione e distanza di una fonte di cibo che ha trovato.
Altri insetti sociali molto noti sono le formiche. La loro organizzazione è simile a quella delle api. Le formiche sono onnivore. Le operaie sono prive di ali, e anche la regina se le strappa dopo il volo nuziale. Vi sono specie di formiche che coltivano funghi nei formicai, altre che accudiscono piccoli insetti, gli afidi, per ricavarne una secrezione zuccherina che “mungono”. Vi sono pure formiche schiaviste che rapiscono larve di altre specie di formiche e utilizzano come operaie gli adulti che si sviluppano da esse.
Sono abbastanza note anche le termiti, che si nutrono di legno e costruiscono nidi con la consistenza del cemento.
Una “sfida” al concetto di selezione naturale
Viene chiamato selezione naturale il processo, determinato dall’ambiente, per cui individui con caratteristiche diverse hanno un diverso successo riproduttivo. In questo modo, le caratteristiche ereditarie favorite dalla selezione naturale si conservano o, se inizialmente rare, divengono sempre più comuni nella popolazione. Se effettivamente la selezione naturale ha un ruolo essenziale nell’evoluzione, si deve aspettare che proprietà importanti delle specie viventi, o di gruppi di livello superiore, siano spiegabili come adattamenti che aumentano il successo riproduttivo di chi le possiede, o almeno che non siano dannose in questo senso.
Gli insetti sociali mostrano una caratteristica che, di primo acchito, sembra incompatibile con il ragionamento precedente. Infatti, le femmine sono predisposte a “rinunciare” alla riproduzione se si sviluppano in celle normali e sono esposte al feromone della regina. Tale caratteristica ereditaria porta, nella stragrande maggioranza dei casi, ad un successo riproduttivo nullo. Sappiamo inoltre che gli insetti sociali discendono da specie solitarie, in cui ogni individuo sviluppa una sua capacità riproduttiva. Lo sviluppo di tali comportamenti sociali rispecchia un caso particolarmente eclatante di altruismo, ossia di comportamento che riduce il successo riproduttivo di chi li mette in atto, a vantaggio di cospecifici.
In effetti un organismo, tranne che nel caso della riproduzione asessuale, non produce individui identici a sé. La forma più frequente di riproduzione sessuale, quella anfigonica, implica che ognuno dei genitori dia ad ogni figlio metà dei propri geni. Il nuovo nato riceve metà del proprio patrimonio ereditario dal padre e metà dalla madre. Ogni figlio fornisce continuità a metà del patrimonio ereditario del genitore per cui, se un individuo ha due figli che sopravvivono è come se egli fosse ancora in vita[non chiaro]. Queste caratteristiche dell’individuo che si è riprodotto, nelle generazioni successive si rimescolano e si diluiscono nella popolazione, quelli che mantengono continuità, salvo mutazioni, sono i geni. Un organismo non è però l’unico possessore dei suoi geni: quelli che ha ricevuto dai genitori saranno stati dati in parte anche a fratelli e sorelle. In media, due fratelli hanno 1/2 dei geni in comune, proprio come genitore e figlio. Tra zio e nipote i geni in comune saranno 1/4, tra cugini 1/8 etc. Ne consegue che, dal punto di vista della sopravvivenza dei geni, avere un figlio o un fratello è esattamente la stessa cosa, avere due nipoti è lo stesso che avere un figlio, e così via.
La rinuncia a prolificare, per permettere ad uno stretto consanguineo di avere molti discendenti, è quindi una “strategia” premiata dalla selezione naturale se la quantità di propri geni che un individuo trasmette “indirettamente” è maggiore di quella che trasmetterebbe riproducendosi da sé. Nel caso delle api, la regina che si riproduce è madre o sorella delle operaie che la aiutano, e produce molti più discendenti di quanti potrebbe produrne un’ape solitaria.
Occorre anche considerare che tutti gli insetti sociali, ad eccezione delle termiti, appartengo all’ordine degli Imenotteri. In tale gruppo, i maschi vengono prodotti per partenogenesi da uova non fecondate. Come risultato, i maschi sono aploidi, e producono spermatozoi che contengono tutti i loro geni, e sono quindi tutti uguali tra loro (questo comportamento, comune solo a imenotteri e pochi altri insetti, si chiama aplodiploide).
Due api femmine con lo stesso padre, hanno la metà del genoma di origine paterna identica e la metà di origine materna per metà uguale e per metà diversa. Il risultato è che due sorelle hanno 3/4 di geni in comune, mentre madre e figlia ne hanno solo 1/2. All’operaia risulta quindi più “conveniente” farsi generare sorelle dalla regina che fare figlie “in proprio”.
Nel caso delle termiti, che non sono aplodiploidi, il ragionamento è diverso: all’interno di una colonia re e regina sono solitamente fratello e sorella. Infatti, quando uno o entrambi i membri della famiglia reale muoiono, il loro ruolo viene preso da due operaie (sia maschi che femmine, al contrario degli imenotteri). In questo caso, soprattutto nei nidi più vecchi, il livello di parentela fra due qualsiasi operaie può arrivare al 100% dei geni. Ancora una volta, è più vantaggioso accudire fratelli che figli.
Bibliografia
Maurice Maeterlinck, La vita delle api (1901), La vita delle termiti (1926), La vita delle formiche (1930): saggi poetici e divulgativi sugli insetti sociali
FONTE
Testo del 10.08.2014
Buona sera amici apicoltori!
Ho rimesso insieme vari appunti presi tempo fa e, come da accordi con Piergiorgio, ho realizzato un breve testo a sua firma per pubblicizzare l’iniziativa in oggetto.
L’idea è la seguente: apportate le eventuali correzioni/modifiche all’articolo, lo pubblicheremo su un prossimo numero di Api&Flora (allegato il testo in word e l’immagine in buona risoluzione) e, parallelamente, sarà mia cura stampare un centinaio di copie in formato A4 da distribuire ai partecipanti del corso 2014/5.
Cosa ne dite? Attendo riscontri,
un abbraccio,
Fabrizio
COME COMUNICANO LE API?
UNA NUOVA INIZIATIVA DI RICERCA SCIENTIFICA E DI AGGREGAZIONE PER I SOCI DELLA C.A.P.T.
Questo annuncio per presentarvi l’avvio di una nuova iniziativa culturale sociale, su iniziativa di Fabrizio e Arianna Urbano, che ci auguriamo riceva il meritato riscontro, coinvolgendo quanti più apicoltori professionisti, amatoriali e neofiti possibile.
L’iniziativa consiste nella creazione di un gruppo permanente di studio che approfondisca interessanti aspetti del mondo apistico, solo apparentemente lontani dall’attività pratica quotidiana degli apicoltori, volta alla produzione del miele e degli altri prodotti dell’arnia: nello specifico si vorrebbero raccogliere informazioni, esperienze e materiale scientifico sui sistemi di comunicazione delle api. Intendendo con queste parole sia lo studio della comunicazione tra api tramite contatto fisico, danze aeree, chimica dei feromoni, termoregolazione dei favi e distribuzione di acqua/alimenti che la comparazione costruttiva con i sistemi di comunicazione delle termiti e degli altri imenotteri sociali (vespe e formiche).
Il Gruppo di Studio sarà basato sul valore fondante la nostra Consociazione, trasmessoci da Don Angeleri, da Carlo Vidano e da tutti gli altri soci che si sono distinti in questi anni per impegno e dedizione: ovvero la passione gratuita e disinteressata per l’apicoltura combinata al sincero amore per le nostre piccole amiche. Pertanto l’iniziativa non avrà alcun risvolto economico: eventuali pubblicazioni che potrebbero derivare saranno di esclusiva proprietà della C.A.P.T. e gli (improbabili) introiti saranno utilizzati per proseguire l’attività statutaria di formazione e diffusione di una cultura apistica responsabile tra le nuove generazioni.
Il Gruppo di Studio è aperto a tutti (apicoltori esperti, dottori e studiosi della materia, dilettanti appassionati e apprendisti stregoni), si incontrerà periodicamente tre-sei volte l’anno nelle sedi istituzionali della C.A.P.T (ovvero Reaglie e Cascina Brero/Cascina Rubbianetta nel Parco della Mandria) e sfrutterà ampiamente una piattaforma informatica dedicata (creata con gli strumenti freeware offerti da Google Inc.) capace di far condividere materiale e opinioni senza limiti di distanze e orari tramite mail, chat e archivi virtuali per documenti-foto-video-mappe. Ovviamente si cercherà di dare supporto ai soci informaticamente meno evoluti ma quanto mai preziosi per l’esperienza maturata sul campo.
Lo studio sui sistemi di comunicazione nell’arnia, e quindi sulle reti neurali che uniscono i singoli soggetti dell’alveare concretando un pensiero comune condiviso, potrà poi essere sviluppato in un secondo momento verso molteplici direzioni, che definiremo insieme proseguendo i lavori. In linea di massima, comunque, queste saranno riconducibili alle reti neurali biologiche (con riflessioni su telepatia, percezioni extrasensoriali, comunicazione tra cellule, etc.) e alle reti neurali artificiali (con riflessioni su informatica e telecomunicazione future come sistemi di controllo di veicoli, simulatori di processi decisionali, riconoscimento di gesti/voci/OCR, diagnosi medica, applicazioni finanziarie, sismologiche e meteorologiche, etc. sino ai c.d. “computer biologici).
Per offrire la propria disponibilità a partecipare al Gruppo di Studio o, semplicemente, per informazioni, scrivete una mail a me o a Fabrizio Urbano.
Testo del 06.09.2014
E’ andata! Il Direttivo della CAPT ha recepito la mia proposta con il consenso di tutti i presenti… prossimamente organizzeremo il primo incontro per tracciare le linee guida operative del gruppo di studio. Ottimo! Mi fa proprio piacere! 😀😀😀
Testo del 30.09.2014
Ho scritto la bozza della mail di presentazione del progetto:
Buona giornata a tutti voi e grazie ancora per aver risposto al nostro appello!
Ottenuta, il 6 settembre scorso, l’autorizzazione dal Direttivo della C.A.P.T per la creazione di un gruppo permanente di studio in seno alla consociazione, è tempo di passare alla fase operativa:
1) Tracciando un primo schema delle linee guida che seguiremo per organizzare i lavori e
2) Fissando la data del primo incontro.
Chi può partecipare al GPS?
Tutti: apicoltori professionisti, amatoriali, neofiti e cultori della materia a condizione di essere validamente iscritti alla C.A.P.T, così da fugare ogni eventuale problematica inerente coperture assicurative legate alla presenza nei locali associativi (ex T.U.S.L. n. 81/2008) e trattamento dei dati riservati dei soggetti partecipanti (ex D.LG. n. 196/2003).
Dove ci riuniremo?
Decideremo di volta in volta tra le tre sedi della C.A.P.T. deputate alla formazione dei soci: nel Parco della Mandria a Cascina Rubbianetta (Centro Internazionale del Cavallo) e a Cascina Brero (Apiario Sperimentale/AL.BO.) e nella sede storica in Strada del Cresto, 2 a Reaglie.
Quando?
Partendo dal presupposto che tutti lamentiamo carenza di tempo, essendo in mille faccende affaccendati, ci incontreremo indicativamente tra le tre e le sei volte l’anno, dando ampio preavviso e preferendo le date già stabilite per altri eventi sociali, così da ottimizzare costi/tempi relativi a spostamenti e riscaldamento/pulizia delle sale utilizzate per riunirci.
Proponiamo per il primo incontro l’xx ottobre 2014 alle ore xx.00 nella Sala di Cascina Rubbianetta (utilizzata la mattina per la lezione del corso di apicoltura 2014/15): siete pregati di confermare la partecipazione entro e non oltre il xx/10/2014 in modo da poter comunicare a tutti il giorno successivo l’ufficializzazione dell’incontro.
Quali obiettivi ci poniamo?
L’acronimo GPS richiama alla memoria il ben più famoso “Global Position System”, il sistema di posizionamento e navigazione satellitare civile che sempre più guida il nostro muoverci sul pianeta… parallelamente il Gruppo Permanente di Studio C.A.P.T. ambisce a divenire uno strumento di guida e indirizzo verso un’apicoltura consapevole e responsabile secondo i valori trasmessici da Don Angeleri, Carlo Vidano e tanti altri soci.
Il progetto si pone principalmente tre obiettivi:
1) Consolidare i rapporti umani tra appassionati di apicoltura;
2) Trasmette informazioni tecniche, esperienze e stimoli di approfondimento scientifico tra consociati;
3) Produrre, eventualmente, brevi dispense riassuntive che, dopo essere state visionate dai nostri professori universitari e da altre alte professionalità del settore, potranno poi essere condivise con altri consociati e ricercatori universitari.
N.B.: Evidenziamo sin d’ora che l’iniziativa non avrà mai alcun fine di lucro. Eventuali pubblicazioni che potrebbero derivare saranno di esclusiva proprietà della C.A.P.T. e gli (improbabili) introiti saranno comunque utilizzati per proseguire l’attività statuaria di formazione e diffusione di una cultura apistica responsabile tra le nuove generazioni.
Quali argomenti approfondiremo?
Ogni incontro durerà tra le due e le quattro ore, durante le quali approfondiremo aspetti pratici e teorici del mondo delle api e dell’apicoltura.
– Per quanto riguarda la “TEORIA”, l’indagine verterà inizialmente sui diversi sistemi di comunicazione tra api: cinetica, chimica, termica ed elettromagnetica oltre alla comparazione con quelli utilizzati dagli altri insetti sociali (es. termiti, vespe e formiche);
– Per quanto riguarda la “PRATICA”, saranno approfonditi aspetti tecnici, patrimonio dell’esperienza degli apicoltori più smaliziati, utili alla miglior gestione delle nostre arnie (es. come costruirsi una verandina o come proteggere le arnie dagli assalti delle formiche).
Come si svolgeranno gli incontri?
Di volta in volta decideremo l’argomento da trattare l’incontro successivo e prepareremo individualmente materiale da presentare agli altri partecipanti: testi da suggerire, foto, video, memorie e quant’altro.
Durante l’incontro, scelto un moderatore, a turno tutti avranno la possibilità di fare propri interventi o relazionare argomenti sui quali si sentono particolarmente ferrati.
Occasionalmente potremo anche invitare ospiti che vorranno trasmetterci (a titolo gratuito) un po’ del loro sapere in determinate materie.
Prima o dopo l’incontro cercheremo di organizzare un frugale buffet nel quale, sapendo in anticipo quanti saranno i partecipanti all’incontro, ognuno potrà liberamente portare qualcosa da mangiare e/o bere all’insegna della serena convivialità, permettendo così la socializzazione tra i diversi soci partecipanti. Terminato il convivio, insieme, riordineremo velocemente la sala.
Quale strumento utilizzeremo per comunicare e condividere materiale tra un incontro e il successivo?
Partiremo molto semplicemente: tutte le comunicazioni avverranno esclusivamente via email.
Ricordo di avere sempre l’accortezza di includere gli indirizzi di tutti i partecipanti in ogni mail, oltre all’indirizzo ufficiale della C.A.P.T., così da permetterci di creare un archivio storico automatico.
Vi suggerisco di inserire i nominativi dei partecipanti nella vostra rubrica e, magari, riunirli in un gruppo di posta elettronica (es. GPS-CAPT) così da velocizzare l’invio. Ad oggi sono:
…
Per quanto riguarda file, testi, video, foto, etc. (copyfree o di cui si detengono i diritti di copyright) che vorrete condividere… in questa prima fase non avrete che da inviare link o allegati all’indirizzo della C.A.P.T. e sarà nostra premura caricarli prontamente in un disco virtuale visibile solo ai soggetti che sono in possesso del seguente indirizzo (che vi suggerisco di salvare nei “Preferiti Internet” del vostro browser): …
Progressivamente, in relazione all’effettivo utilizzo degli strumenti da parte dei partecipanti al GPS-CAPT, potremo attivare il caricamento multimediale autonomo, la chat testuale/audio/video, il social network Google+, strumenti office condivisi, etc.
Ma non mettiamo troppe pentole sul fuoco che poi rischiamo di bruciarle! Piano piano facciamo tutto!
A presto,
Fabrizio Urbano
Buona giornata a tutti!
– Lo scorso sabato 11 ottobre 2014 alle 11:00
– Presso i locali di Cascina Rubbianetta, nel Parco Regionale della Mandria
– Fabrizio, Arianna, Piergiorgio, Filippo, Gabriele, Elena, Matteo, Giuseppe (e Giancarlo)
Si sono riuniti per la prima volta al fine di definire le linee guida che indirizzeranno l’operatività del Gruppo di Studio.
Si è deciso che:
1) Il prossimo incontro si terrà nello stesso luogo alle ore 10.30 di sabato 15 novembre 2014;
2) Per quanto riguarda la ricerca “TEORICA”, tutti i partecipanti rileggeranno il capitolo dedicato alla “Vita sociale delle api” del testo di Giovanni Bosca “Guida pratica di apicoltura”, Ed. Il Castello, qualunque edizione;
3) E tutti i partecipanti raccoglieranno materiale (libri, riviste, articoli, memorie, etc.) inerenti l’argomento “I sistemi di comunicazione tra gli insetti sociali” con una particolare attenzione alla comunicazione elettromagnetica-neuronale (meno esplorata rispetto a quella chimica e cinetica);
4) Per quanto riguarda la ricerca “PRATICA”, rimandiamo l’argomento da approfondire al primo incontro del 2015 poiché prima il Presidente vuole “rodare” il sistema di acquisizione e condivisione delle fonti per le ricerche teoriche.
5) In seno al Gruppo GPS-CAPT, il Presidente ha chiesto ad alcuni soggetti (Fabrizio, Giancarlo, Filippo, Antonio) di costituirsi in sottogruppo temporaneo per aggiornare i riferimenti (indirizzi, mail, tel., etc.) obsoleti presenti sulla tessera CAPT e su Api&Flora oltre al produrre alcune proposte per ringiovanire le grafiche da sottoporre al Direttivo del 13 dicembre p.v. >>> Seguirà mail specifica ai soggetti interessati, ma se altri vogliono partecipare sono i benvenuti!
A presto,
Fabrizio Urbano
innanzitutto voglio sinceramente ringraziare tutte le persone che hanno risposto all’appello pubblicato sull’ultimo numero di Api&Flora. In particolar modo chi, nonostante il tempaccio che imperversa sul nostro territorio, stamane ci ha raggiunti a Cascina Rubbianetta per delineare insieme l’operatività futura del Gruppo di Studio Permanente della C.A.P.T..A seguire il report per punti di quanto è stato relazionato e deciso per il futuro prossimo:1) Hanno risposto all’appello una ventina di membri della Consociazione… equamente divisi tra matricole volenterose di conoscere più in profondità il mondo dell’apicoltura e maestri desiderosi di condividere la propria esperienza.
Nello specifico il GPS ad oggi è composto da: A. Barbiso, A. Massari, A. Pautasso, B.F. Ciriano, E. Magnani, F. Urbano, F. Giordano, G. Colombano, G. Benedetto, G. Migliore, G. Filippa, J.G. Barriga, M. Pinto, M. Zanghi, P. Bonci, S. Suraci, S. Piantedosi. Più diversi altri “osservatori”.
Se volete aggiornare i vostri “gruppo di invio mail”, potete aggiungere:
Benito Franco Ciriano e Giovanni Migliore.2) Alcuni partecipanti hanno avuto difficoltà nel trovare il luogo d’incontro.
Per evitare problemi futuri, ricordo che abbiamo realizzato una mappa su Google Map con indicata la posizione di tutte le sedi e di tutti i fornitori di materiale apistico della C.A.P.T..
La mappa è raggiungibile CLICCANDO QUI
Vi suggerisco di salvare il link nei vostri preferiti Web e vi ricordo che la mappa è interattiva: ruotando la rotellina del mouse è zoomabile e, inserendo la vostra posizione, otterrete il percorso più rapido per raggiungere la meta.
Se utilizzate una periferica Android e la connessione satellitare, si attiverà in automatico anche il navigatore gps.3) Per quanto riguarda lo studio di fattibilità di un’analisi sulla comunicazione elettromagnetica tra le api, i membri del GPS hanno raccolto materiale attinente la materia (comunicazione elettrica dei fiori verso le api, interferenza di linee elettriche e reti wi-fii/cell sulle api, comunicazione cinetica e chimica tra le api, …) ma nulla di specifico.
Ciò rende ancora più utile e interessante l’approfondimento rinnovando la comune volontà di ricerca.
In particolar modo concentreremo le prossime ricerche approfondendo gli studi sulla “presenza di magnetite nel cervello di api e altri animali…” elemento fondamentale per l’instaurarsi di campi elettromagnetici.4) Sulla base dei suggerimenti ricevuti ho riorganizzato il disco virtuale condiviso nel quale raccoglieremo i frutti delle nostre ricerche. Cliccando su questo link, che vi suggerisco ancora di salvarvi nei preferiti Web ( DISCO VIRTUALE DEDICATO ) potrete accedere a tre cartelle nelle quali sono raccolti:
– l’Archivio delle mail inviate;
– l’Archivio delle immagini (foto e disegni) che vogliamo conservare;
– l’Archivio dei testi che vogliamo condividere.
NB: se non avete esperienza nel salvare documenti online in formato pdf… semplicemente scrivetemi a questo indirizzo una mail con inserito il link alla/e pagina/e Web che volete condividere e poi ci penserò io.
5) Mentre ci siamo… cerchiamo di fare un nuovo piccolo passo nell’utilizzo dei programmi accessori del pacchetto Google.
Nel disco condiviso, oltre alle tre cartelle descritte al punto 4, troverete un file word dal titolo “Bibliografia consigliata”.
Cliccandoci sopra una volta, potrete aprirlo e leggerlo.
Se passate il puntatore del mouse sulla parte superiore del documento… scenderà una tendina nera, nel centro della quale vedrete il pulsante “APRI”. Cliccandolo, il documento si aprirà nel programma di scrittura online e potrete integrarlo personalmente. Il programma permette gratuitamente di scrivere contemporaneamente a sedici persone… credo ci basti e avanzi!
6) Abbiamo chiesto a Bonci di procurarci un elenco, anche cartaceo, dei libri presenti nella biblioteca della Consociazione a Reaglie, così da poter cercare sinossi online e poter evidenziare testi utili alla nostra ricerca.
Nel caso in cui il documento non fosse fruibile… potremmo organizzarci un giorno per visionare i testi personalmente nella sede in collina… magari mangiando poi un boccone insieme nella vicina trattoria.
7) Il sito ufficiale dell’Accademia Nazionale di Entomologia è raggiungibile cliccando: QUI
Nella sezione dedicata ai rendiconti si trovano moltissimi documenti utili e certificati.
8) Per concludere… il prossimo incontro del GPS-CAPT si terrà sabato 17 o 24 gennaio 2015, verosimilmente nella sala adiacente l’apiario didattico in Cascina Brero-ALBO.
Indicativamente, la prima parte dell’incontro sarà dedicata all’approfondimento di una qualche attività manuale sulle arnie a cura di Bonci e la seconda alla relazione di quanto approfondito in materia di comunicazione tra le api.
Seguirà comunque mail specifica d’invito intorno al 10/1 p.v. con orario, luogo e argomenti specifici.
A presto,
Fabrizio
Per evitare coinvolgimenti istituzionali e possibili fastidi da parte di soggetti non graditi (leggi “troll” e “spammer”), in questa prima fase di test amministrerò il gruppo e la richiesta di iscrizione sarà sottoposta ad approvazione.
Condividete e partecipate numerosi!
LINK PER GRUPPO FACEBOOK
LINK PER COMMUNITY GOOGLE PLUS: (NON PIU’ ATTIVO DAL 02.04.2019 PER CHIUSURA SOCIAL NETWORK DA PARTE DI GOOGLE)


{kind=link}
{kind=link}
{kind=link}